在线粒体中,肌动蛋白调节线粒体DNA转录,是线粒体质量控制所必需的
作者:Xin Xie,1 Tomas Venit,1 Nizar Drou,2 and Piergiorgio Percipalle1,3,4,*
总述:
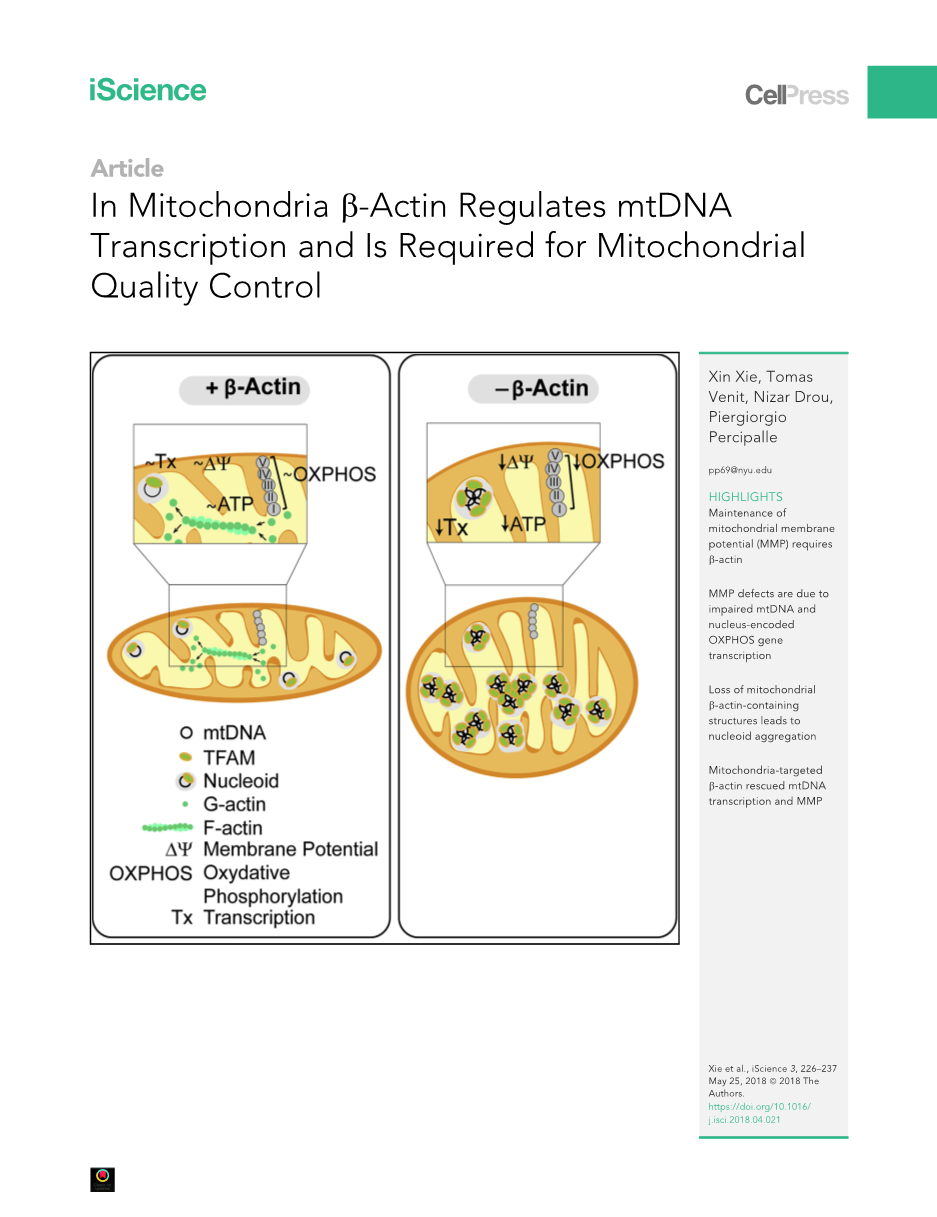
在真核细胞中,肌动蛋白同时调节细胞质和细胞核的功能。然而,线粒体中是否存在以肌动蛋白为基础的结构并参与线粒体的功能尚未得到研究。使用野生型肌动蛋白 / 和敲除(KO)b-肌动蛋白小鼠胚胎成纤维细胞,证明了b-肌动蛋白缺失细胞在维持线粒体膜电位(MMP)方面具有缺陷。MMP缺陷与线粒体DNA(MtDNA)转录和核氧化磷酸化(OXPHOS)基因表达受损有关。使用超分辨率显微镜,我们提供了线粒体内含有b-肌动蛋白结构的直接证据。在KO细胞的球状线粒体中观察到大量的TFAM染色的类核聚集体,提示线粒体类核分离存在缺陷,缺乏b-actin。线粒体靶向b-actin挽救mtDNA转录和MMP的观察表明,线粒体b-actin池对于线粒体质量控制具有不可或缺的功能作用。
引入:
肌动蛋白在细胞质中调节细胞形态、运动、细胞器动态和功能(Dominguez和Holmes,2011)。已知胞浆肌动蛋白与线粒体的特定相互作用介导了线粒体网络的分裂和线粒体的运输,有助于线粒体的细胞分布(Boldogh和Pon,2006;Senning和Marcus,2010)。尽管确切的机制仍未完全揭开,胞浆肌动蛋白也参与了线粒体依赖性的细胞凋亡(Gourlay和Ayscough,2005)。证据还表明,肌动蛋白和一些肌动蛋白相关蛋白,如肌球蛋白,参与了线粒体供能与线粒体DNA(MtDNA)的关联(Reyes等人,2011年)。
在原核生物和真核生物中都描述了肌动蛋白与DNA的功能联系(MollerJensen等人,2002年;Visa和Percipalle,2010年)。在真核细胞核中,肌动蛋白与活性基因相互作用,并通过所有三种真核细胞核RNA聚合酶控制转录(Visa和Percipalle,2010)。在基因组水平上,b-actin调节染色质的分布和表观遗传标记的沉积,导致基因程序的激活或抑制,并影响细胞身份(谢等,2018年)。在杆状细菌中,肌动蛋白样蛋白在基因组和质粒DNA分离中起着重要作用(Kruse和Gerdes,2005;Moller-Jensen等人,2002)。考虑到它们的环状基因组,以及线粒体DNA的维持和复制是独立执行的事实,基于肌动蛋白的机制可能是线粒体DNA分离的基础。事实上,b-actin和肌球蛋白被发现与线粒体类核有关,并且在分离的线粒体中发现了一个抵抗蛋白酶消化的b-actin池(Reyes等人,2011年),这表明b-actin定位于线粒体中。然而,线粒体内的肌动蛋白是如何组织的,线粒体肌动蛋白是否在线粒体质量控制中发挥作用尚不清楚。
在这项研究中,我们分析了野生型(WT)b-actin / 小鼠和基因敲除(KO)b-actin-/-小鼠的胚胎成纤维细胞(Tondeleir等人,2012年)。当比较缺乏功能性肌动蛋白等位基因的KO细胞和WT细胞时,我们发现在维持线粒体膜电位(MMP)方面存在严重缺陷。这种缺陷可以归因于线粒体DNA转录受损和核编码的氧化磷酸化(OXPHOS)基因下调。使用超分辨显微镜,我们观察到线粒体内含有b-肌动蛋白的结构,这些结构似乎与胞浆中的对应结构相连。在缺乏B-肌动蛋白的情况下,线粒体类核倾向于在球状线粒体中形成大的聚集体。再加上KO细胞中mtDNA拷贝数的显著增加,我们的发现表明线粒体DNA复制后,线粒体类核出现分离缺陷。重要的是,线粒体靶向b-肌动蛋白在组成性地重新导入b-肌动蛋白缺失细胞时,挽救了mtDNA转录和基质金属蛋白酶。总体而言,我们的研究提供了线粒体中存在B-肌动蛋白的直接证据,并揭示了线粒体池中B-肌动蛋白在线粒体质量控制中的未知作用。
图1 b-actin基因敲除中线粒体膜电位受损和对线粒体应激的超敏反应
结果:
- 缺失b-actin的细胞线粒体膜电位(MMP)维持缺陷
我们利用高含量筛选平台分析了WT(b-actin / )和KO(b-actin-/-)小鼠胚胎成纤维细胞的线粒体特征。E m b r y o s(Tondeleir等人,2012年)。用MitoTracker橙色染料染色的细胞,根据膜电位在线粒体中积累,固定后保留良好,对单个细胞中染色的线粒体斑点(MitoTracker橙色阳性染色)进行定量分析(图1A)。我们发现,与WT相比,KO细胞质内有较多染色斑点(图1B)。然而,KO细胞的平均光斑强度明显较低,平均光斑面积较小(图1C和1D)。平均而言,KO细胞每个细胞的总斑点面积和强度值也明显低于KO细胞(图1E和1F)。这些数据表明,在没有B-肌动蛋白的情况下,较小的线粒体区可以维持膜电位,导致KO细胞的MMP总体较低。共聚焦图像基本上证实了高含量筛选的结果(图1G)。荧光激活细胞分选(FACS)分析显示,基于MitoTracker橙色染色,KO细胞丢失了超过50%的MMP(图1H)。在只有一个功能性b-actin等位基因的杂合子细胞中,未观察到MMP的显著丢失(图S1a)。因此,只有在缺少两个功能性肌动蛋白等位基因的情况下,才会发生严重的MMP丢失。
我们进一步使用四甲基罗丹明乙酯(TMRE)对活细胞进行染色,TMRE依靠基质金属蛋白酶(MMP)动态积累或与线粒体结合。TMRE对WT和KO细胞的染色强度随TMRE浓度的降低而减弱。然而,在所有浓度下,KO细胞的TMRE染色强度明显低于WT细胞(图1I)。值得注意的是,在较低浓度下,WT和KO细胞的TMRE强度差异变大(图1I)。因此,KO细胞在积累TMRE方面存在缺陷,尤其是在低浓度时,表明维持动态MMP的能力受损。
- b-肌动蛋白缺失时线粒体应激敏感性增加和线粒体形态改变
接下来,我们研究了MPP (1-甲基-4-苯基吡啶,一种特异性复合物I抑制剂)和CCCP(羰基氰化物间氯苯腙,一种线粒体解偶联剂)诱导的线粒体应激下线粒体膜电位的变化(齐等,2013年)。MPP 和CCCP处理减少了WT和KO细胞的TMRE染色(图1J,左图)。然而,在MPP 和CCCP应激处理后,KO细胞失去了更多的MMP(TMRE强度)(图1J,右图)。一直以来,活细胞图像显示KO细胞对CCCP压力非常敏感,并且在加入CCCP后细胞迅速萎缩(视频S1)。此外,与WT细胞相比,KO细胞在高浓度CCCP去极化线粒体膜后,线粒体膜MMP的恢复也受到抑制(图S1B和S1C)。综上所述,这些数据表明,缺乏b-actin的细胞对线粒体膜损伤引起的线粒体压力更敏感,而线粒体膜损伤又与线粒体膜维持的内在缺陷有关。
我们进一步应用电子传递链复合物的选择性抑制剂,发现KO细胞对复合物I、II和III的活性抑制更敏感,表现为与WT细胞相比,MMP的损失更大(图1K和1L)。重要的是,DCCD(N,N-二环己基碳二亚胺)抑制复合体V(ATP合成酶)导致WT和KO细胞的MMP增加;然而,KO细胞中MMP的增加程度要高得多(图1L)。增加了细胞的线粒体膜电位。由于DCCD抑制ATP合成酶(复合物V)中的质子转移而对电子转移活性没有显著影响(Clejan等,1984;Toei)。
我们的结果表明,当ATP合成酶被阻断时,KO细胞的线粒体膜间间隙比WT细胞有更多的未使用的质子储存容量。因此,KO细胞的线粒体膜电位降低是由于线粒体缺乏质子积累,而不是由于线粒体缺乏质子储存能力。
几项研究表明,MMP的改变可能导致线粒体形态的改变(Leonard等人,2015年;Safiulina等人,2006年)。因此,我们使用MitoTracker Deep Red研究了活细胞中线粒体的形态,它在线粒体中积累,而不考虑基质金属蛋白酶(MMP)。不出所料,我们发现大多数WT细胞显示出细丝状线粒体网络。相反,KO细胞线粒体肿胀,呈球状(图S1d和视频S2)。由于线粒体肿胀和体积增大与MMP的丢失有关(Leonard等人,2015年;Safiulina等人,2006年),KO细胞中观察到的线粒体形态很可能是由严重受损的MMP引起的。我们进一步测试了减少WT细胞中的MMP是否会导致类似的形态学变化。在MPP 存在下,我们观察到WT细胞经30min处理后逐渐形成球状线粒体(图S1E)。活细胞图像显示,最初的杆状线粒体沿长度逐渐缩短,变成球形(视频S3)。经MPP 处理后,KO细胞的球状线粒体水平也明显增加(视频S3)。总而言之,我们的数据表明,b-actin在控制MMP和线粒体形态方面是不可或缺的。
图2 b-actin基因敲除中,OXPHOS缺陷与线粒体DNA和核OXPHOS基因表达受损有关
- 在b-肌动蛋白敲除中,线粒体DNA和细胞核中OXPHOS基因转录受损与OXPHOS活性降低有关
在OXPHOS期间,MMP由电子传输链复合物的活性维持(Dzbek和Korzeniewski,2008)。为了研究在没有B-肌动蛋白的情况下电子传递活性,我们分离了WT和KO细胞的线粒体,并比较了复合体II/III的活性。KO细胞的线粒体显示复合体II/III活性显著降低(图2A)。一致地,KO细胞产生比WT细胞更低的细胞ATP水平(图2B),在没有B-肌动蛋白的情况下,共同显示出OXPHOS活性受损。然后,我们使用最近发布的RNA-SEQ数据集分析了核OXPHOS基因的相对表达水平(谢等人,2018年)。值得注意的是,核OXPHOS基因在WT和KO细胞的差异表达(DE)基因中显著过表达(图2C)。这些结果表明,KO细胞中OXPHOS活性受损是OXPHOS基因表达改变的直接结果。与受损的 MMP 一致,大多数 DE OXPHOS 基因在 KO 细胞中显示下调(图 2D)。这些下调的基因主要编码电子传递链中复合物I、II和III的组成部分(图2e)。我们通过定量聚合酶链反应(QPCR)证实了Ndufs3、SDHA和Uqcrbin KO细胞的低表达(图2F)。OXPHOS基因的整体下调趋势与先前的发现一致,即OXPHOS的核成分在转录上是共同调控的(van Waveren和Moraes,2008)。
电子传递复合体的基本成分也由线粒体基因组编码。有明显的TB-actin和myosin在mtDNA拓扑和拷贝数维持中发挥作用(Reyes等人,2011年)。然而,b-actin如何潜在地调节mtDNA拷贝数和mtDNA转录尚不清楚。因此,我们分析了KO细胞线粒体DNA拷贝数及其转录本的状况。我们发现,在没有B-肌动蛋白的情况下,线粒体DNA拷贝数增加,这是通过量化mtDNA基因座的子集来揭示的(图2G)。此外,总DNA测序还显示KO细胞mtDNA基因组的mtDNA水平总体较高(图S2A)。然而,线粒体DNA转录本在KO细胞中的表达显著降低(图2H)。线粒体DNA转录本水平的降低可能是由于RNA合成减少或RNA降解增加所致。为了区分这两种可能性,用5-乙炔基尿苷标记新合成的RNA,以监测RNA合成和衰变的速率(图2i,另见透明方法)。对于RNA合成,我们对细胞进行了2小时和4小时的脉冲标记,以量化2小时间隔内相对RNA的变化(图2I,左面板)。在RNA衰变实验中,标记细胞在新鲜培养基中培养,以确定20小时后标记RNA的剩余量(图2I,右侧面板)。我们观察到KO细胞中b-actin的缺失会导致RNA合成受损,而不会影响RNA的衰变(图2ja和d2K)。总之,我们的数据表明,在缺乏B-actin的情况下,核OXPHOS基因下调,mtDNA转录受损,这表明核基因组和线粒体基因组协调一致,以平衡OXPHOS组分的表达。OXPHOS亚单位的核和线粒体表达之间的相关性在以前的研究中也有报道(Gagnon等人,1991;Murdock等人,1999)。与OXPHOS组分表达减少一致,KO细胞通过MitoTracker深红染色评估显示总体线粒体质量较低(图S2B)(Schieke等人,2006年)。
图3 线粒体中有一团b-肌动蛋白,没有b-肌动蛋白池,TFAM染色的核团往往会形成大的聚集体
- 在没有B-肌动蛋白的情况下,线粒体内有一个B-肌动蛋白池,线粒体核团形成大的聚集体
由于核内的OXPHOS基因和线粒体DNA的转录都不受b-actin的影响,因此一个关键的问题是b-actin主要在细胞质还是细胞核内发挥作用来控制OXPHOS基因。我们首先使用了最近发表的含有绿色荧光蛋白(GFP)-b-actin和GFP-核定位信号(NLS)-b-actin的质粒(Sharili等人,2016年),并在KO细胞中瞬时表达了它们。不出所料,GFP-b-actin主要分布在细胞质中,NLS-b-actin主要分布在细胞核中(图S3A)。然而,b-肌动蛋白和NLS-b-肌动蛋白对KO细胞的MMP均无拯救作用(图S3B-S3D)。KO细胞中过表达的外源性b-actin水平远低于WT细胞中的内源性b-actin水平(图S3E)。然而,核靶向b-actin未能挽救MMP的事实促使我们研究b-actin在线粒体中的潜在作用。
先前的一份报告显示,mtDNA与b-actin和肌球蛋白相关,分离的线粒体中的b-actin池似乎对蛋白酶消化有抵抗力(Reyes等人,2011年;Wang和Bogagen,2006年)。然而,没有直接证据表明含有b-肌动蛋白的结构存在于哺乳动物细胞的线粒体中。为了证明
剩余内容已隐藏,支付完成后下载完整资料

英语原文共 31 页,剩余内容已隐藏,支付完成后下载完整资料
资料编号:[597891],资料为PDF文档或Word文档,PDF文档可免费转换为Word
课题毕业论文、文献综述、任务书、外文翻译、程序设计、图纸设计等资料可联系客服协助查找。
您可能感兴趣的文章
- 在线粒体中,肌动蛋白调节线粒体DNA转录,是线粒体质量控制所必需的外文翻译资料
- 水稻转运蛋白——自然抗性相关巨噬细胞蛋白(NRAMP)家族的生物信息学分析外文翻译资料
- 大麦籽粒总淀粉、直链淀粉和支链淀粉的全基因组关联分析揭示新的等位基因外文翻译资料
- 人工合成群落中食细菌线虫与捕食性细菌的行为交互作用外文翻译资料
- RNA靶向的CRISPR系统 ——从宏基因组到RNA调控外文翻译资料
- 通过农杆菌真空浸润和共培养渗透(发芽)种子使烟草脆裂病毒介导小麦和玉米的全株水平病毒诱导的基因沉默外文翻译资料
- 一种来自盐生植物Atriplex Canescens(Pursh)Nutt.的重金属分离蛋白质(AcHMA1),在酿酒酵母中表达对铁和其他非生物胁迫的耐受性外文翻译资料
- 初中生信息素养概况外文翻译资料
- 概念转变:对科学教育的理论,方法和实践挑战的探讨外文翻译资料
- 互联网学习满意度的性别差异外文翻译资料

